|
|
|
|
|
진핵세포는 핵을 비롯한 다양한 기능을 하는 세포소기관을 갖는다. 진핵세포가 세포분열 할 때, 소화작용을 일으키는 세포소기관인 ‘리소좀’의 유전을 조절하는 단백질의 작동원리가 세계 최초로 밝혀져 주목받고 있다. UNIST(총장 이용훈) 생명과학과 이창욱 교수팀은 GIST(총장직무대행 박래길) 생명과학과 전영수 교수팀과 함께 공동연구를 진행했다. 연구팀은 효모 모델을 활용해 세포분열 시 자가포식에 관여하는 리소좀 단백질인 Vac8이 결합파트너 단백질인 Vac17과 복합체를 형성해 자가포식을 억제하고 분열된 세포로 리소좀 수송을 조절한다는 사실을 세계 최초로 밝혀냈다. 이와 관련된 연구는 향후 세포분열 이상으로 발생하는 암, 만성질환 등 치료연구에 있어서 중요한 초석이 될 것으로 전망된다. 세포분열은 인간을 비롯한 생명체의 생명유지에 있어 필수 불가결한 요소이다. 그래서 이는 많은 세포 내 기전들로 인해 정교하게 조절된다. 특히 완전하고 안전한 세포분열을 위해서는 세포 내 자가포식조절이 매우 중요하다. 자가포식 활동으로 분열하는 세포의 세포소기관 및 구성물질 등이 분해되면 정상적인 딸세포로의 분열이 이루어질 수 없기 때문이다. 리소좀 단백질인 Vac8은 여러 결합파트너와 복합체를 형성해 선택적으로 자가포식을 조절한다고 알려져 있다. 또한, Vac17, Myo2 단백질과 결합해 세포분열 시 리소좀 수송에도 관여한다. Vac8이 Vac17, Myo2와 삼중 구조체를 형성하고, 이렇게 형성된 복합체는 일종의 세포 내 도로의 역할을 하는 액틴 단백질을 통해 모세포에서 딸세포로 리소좀을 수송한다는 것이다. 기존 연구를 통해 Vac8과 Vac17 단백질의 복합체 형성이 리소좀 수송에 중요하다는 사실이 밝혀져 있었으나, Vac8이 어떠한 원리로 세포 내 자가포식을 억제하고, 리소좀 수송 기능을 수행할 수 있는지에 대한 분자 수준의 연구는 밝혀지지 않았다. 본 연구팀은 X-ray 구조법을 통해 Vac8-Vac17 복합체의 고해상도 구조를 세계 최초로 규명했다. 해당 복합체는 두 개의 결합 인터페이스를 형성한다. 특히 연구팀은 인터페이스 형성에 직접 관여하는 아미노산을 확인했으며, 이 아미노산에 돌연변이가 일어난 Vac8 및 Vac17을 가진 효모에서는 세포 분열시 리소좀이 딸세포로 이동하지 못하는 것을 관찰했다. Vac8 단백질이 리소좀 수송을 조절한다는 것을 직접적으로 입증한 것이다. 또한 이번 연구에서는 Vac8이 특정 결합 파트너와 복합체를 이뤘을 때 형성하는 4차 구조가 자가포식과 리소좀 수송을 구별해 조절한다는 사실을 입증해냈다. 연구팀은 이러한 구조적 차이가 Vac8에 포함된 ARM 도메인과 N-말단의 H1 helix에 의한 조절임을 밝혀냈다. Vac8-Vac17의 복합체가 형성되면, Vac17이 Vac8의 H1 helix를 붙잡게 되고, H1 helix가 ARM도메인으로부터 분리되지 않게 만들어 자가포식을 일으키는 복합체(Vac8-Nvj1, Vac8-Atg13) 형성을 억제한다. 이는 세포분열 시, 자가포식이 억제되고 리소좀의 수송이 일어난다는 것을 의미한다. 이창욱 생명과학과 교수는 “이 연구는 세포분열 시 자가포식 활동의 억제와 리소좀 수송이 한가지 단백질에 의해 어떻게 조절될 수 있는지를 보여준 중요한 사례다”며 “세포분열과 관련된 질병인 암 및 여러 종류의 만성질환 등의 연구에 대한 중요한 초석이 될 것이다”고 밝혔다. 이번 연구결과는 미국과학학술원회보(PNAS) 4월 24일자로 게재됐다. 연구 수행은 과학기술정보통신부 한국연구재단(선도연구센터지원사업, 중견연구자지원사업, 바이오융합원천기술개발사업) 등의 지원을 받아 이뤄졌다. (논문명: Structures of Vac8-containing protein complexes reveal the underlying mechanism by which Vac8 regulates multiple cellular processes) |
|
|
|
[붙임] 연구결과 개요 |
1. 연구배경생명체의 기본단위인 세포는 여러 기능을 수행하며 끊임없이 분열한다. 이러한 세포분열 시에는 자가포식 작용의 조절이 필요한데, 자가포식 작용이 일어나게 되면, 세포소기관을 비롯한 세포 내 물질들이 분해되어 온전한 딸세포로의 분열이 일어나지 않을 수 있기 때문이다. Vac8 단백질은 리소좀에 존재하는 단백질로써, Nvj1, Atg13과 같은 결합 파트너와 복합체를 이루면, Cvt, PMN과 같은 세포 속 선택적 자가포식 기능을 조절하는 단백질이라고 알려져 있다. 또한, Vac8 단백질은 선택적 자가포식 작용을 조절할 뿐 아니라, Vac17과 Myo2단백질과 결합하여 리소좀의 딸세포로의 이동에도 관여한다고 알려져 있다. 리소좀 단백질인 Vac8이 Vac17을 매개로 Myo2와 결합하게 되면, Myo2는 액틴과 결합하고 이동하여 리소좀을 수송시키는 원리이다. 이러한 세포 내 리소좀 이동에 대한 사실은 알려져 있으나, 어떻게 Vac8과 Vac17이 결합하여 해당 역할을 수행하는지, 그리고 세포분열 시 어떠한 원리로 세포 내 자가포식을 억제하고, 리소좀 수송 기능을 수행할 수 있는지에 대한 분자적 수준의 연구는 아직 미지의 영역으로 남아 있다. |
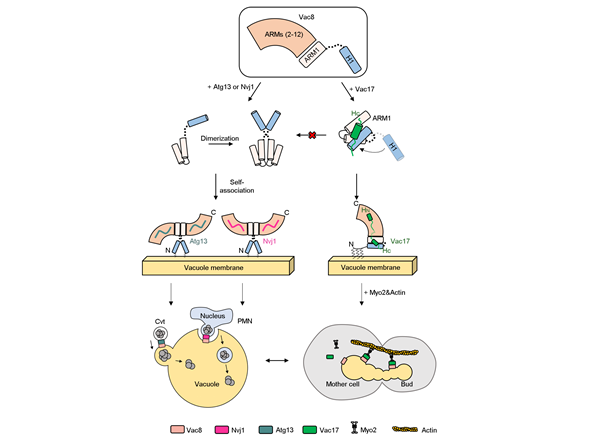
2. 연구내용이번 연구는 구조를 기반으로 세포 분열시 리소좀의 수송에 관련된 Vac8-Vac17 복합체의 작동 원리를 규명하는 것과 해당 기작이 자가포식작용과 어떻게 독립적으로 일어나며 서로를 조절할 수 있는지에 대한 이해를 목표로 두고 진행했다. 연구팀은 X-선 구조 분석법을 사용하여 리소좀 수송에 관여하는 Vac8-Vac17 복합체의 고해상도 구조를 규명했다. 해당 구조에서는 특이하게도, Vac8-Vac17 복합체 사이에는 기존의 Vac8과 결합파트너 구조에서 보인 인터페이스와 유사한 인터페이스 I, 그리고 Vac8-Vac17 복합체에서만 특이적으로 존재하는 인터페이스 II가 존재함을 확인했다. 특히 인터페이스 II는 Vac8의 H1 helix와 직접적인 결합을 통해 형성된다는 특징이 있다. 해당 부분의 결합 인터페이스 형성을 방해하는 돌연변이 제작을 통해 각각 인터페이스가 복합체 형성에 중요한지를 등온 열량 적정계와 풀-다운 실험을 통해 확인해 본 결과, 두 인터페이스가 모두 정상적으로 만들어져야 단백질 복합체가 형성된다는 사실을 확인했다. 또한, 해당 복합체의 형성이 방해되면, 실제 세포 속에서도 세포 분열시 리소좀 수송이 일어나지 않는다는 사실을 직접적으로 입증했다. 또한 본연구에서는 Vac8이 특정 결합 파트너와 복합체를 이루었을 때, Vac8 단백질이 형성하는 사차 구조가 자가포식과 리소좀 수송을 구별하여 조절한다는 사실을 입증해냈다. 이전에 밝혀진 Vac8-Nvj1, Vac8-Atg13 복합체가 이형-사량체를 형성하는 것과 다르게 Vac8-Vac17 복합체는 이형-이량체를 형성하는 것을 단백질의 구조와 크기 배제 크로마토그래피 방식으로 확인할 수 있었다. 본 연구에서 이러한 구조적 차이는 Vac8의 H1 helix에 의해 조절된다는 사실을 밝혀냈다. H1 helix는 ARM도메인 단백질인 Vac8의 N-말단에 존재하는 약 15개의 아미노산으로 이루어져 있다. H1 helix는 ARM도메인과 강하게 결합하지 않고 약한 결합으로 존재한다. 그래서 자가포식을 일으키는 결합파트너인 Nvj1과 Atg13이 Vac8과 결합하게 되면, H1 helix는 ARM도메인으로부터 분리 되어 Vac8이 이량체를 형성하게 된다. 하지만 Vac8-Vac17 복합체가 형성되면, Vac17의 Hc helix가 Vac8의 H1 helix와 결합하여 H1 helix가 ARM1으로 부터 분리될 수 없게 한다. Vac17이 H1 helix가 떨어지지 못하도록 붙잡는 역할을 하는 것이다. 이렇게 Vac8의 이량체 형성이 저해되면, 자가포식을 일으키는 단백질인 Nvj1, Atg13과 Vac8의 복합체 형성을 저해하게 된다. H1 helix의 분리가 Vac17에 의해 억제되면, Vac8은 선택적 자가포식을 일으키는 복합체를 형성할 수 없게 된다. 이는 Vac8이 자가포식을 억제하고 리소좀 수송을 일으키며 세포분열이 가능한 환경을 조성하도록 세포 내 기능을 조절하는 것을 의미한다. |
3. 기대효과세포분열 시 세포는 다양한 메커니즘을 조절함으로써 모세포에서 딸세포로 정상적인 기능을 하는 여러 세포로 나눠진다. 이러한 세포분열에서는 자가포식 작용의 조절이 매우 중요하다. 현재까지는 특히 세포 내 청소작용을 하는 자가포식이 세포분열 시 어떠한 방식으로 억제가 되고, 세포소기관의 유전이 일어나는지에 대한 연구가 부족했다. 이번 연구를 통해, 세포분열 시 리소좀의 수송과 자가포식의 프로세스를 Vac8-Vac17 결합체를 통해 조절할 수 있음을 세계 최초로 입증했다. 해당 연구는 세포분열 시 단일단백질이 어떤 방식으로 세포 내 프로세스를 조절할 수 있는지를 이해시켜주는 중요한 발견이다. 이는 결국 세포의 비정상적 분열로 인해 발생하는 질병인 암, 그리고 여러 종류의 만성질환 등의 치료연구에 있어서 중요한 초석이 될 것이다. |
|
[붙임] 용어설명 |
1. 세포분열 (Mitosis)하나의 모세포가 분열하여 2개 또는 그 이상의 딸세포로 나뉘는 과정을 말한다. |
2. 리소좀 (Lysosome)세포내에서 소화작용을 일으키는 세포소기관. 바이러스와 같은 외부 침입자와 세포내 노폐물 등을 분해하는 역할을 한다. |
3. 자가포식 (Autophagy)자가포식은 세포 속에 노폐물이 쌓이거나 바이러스 같은 외부 침입자가 들어오거나 세포내 에너지가 부족하면 시작된다. 불필요한 물질을 세포 내에서 스스로 분해하여 스스로 에너지를 생산하는 작용이다. |
4. 이형이량체 (Hetero-dimer)서로 다른 두가지의 단백질이 결합 되어 복합체를 형성하는 것을 말한다. |
5. 이형사량체 (Hetero-tetramer)서로 다른 네가지, 혹은 두가지의 다른 이량체 단백질이 결합 되어 네 개의 단백질로 이루어진 복합체를 형성하는 것을 말한다. |
6. ARM도메인 (Armadillo repeat domain)약 42개의 아미노산으로 이루어진 3개의 Helix로 구성된 모티프들의 반복된 도메인으로써, 여러 단백질이 결합할 수 있는 결합 인터페이스의 역할을 한다. |
7. H1 helix (H1 helix)Vac8의 ARM도메인이 시작하기전 N 말단에 존재하는 알파 나선형 구조로써, Vac8의 이량체 형성을 조절한다. |
8. 단백질 4차 구조 (Quaternary structure)단백질 복합체에서 다수의 접힌 단백질 서브 유닛의 배열이다. |
|
[붙임] 그림설명 |
|
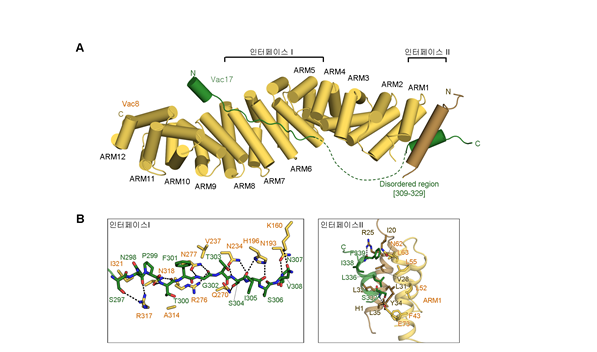
그림1. Vac8-Vac17의 단백질 결정 구조.A. Vac8-Vac17 단백질 복합체 구조. 두 부분의 인터페이스로 복합체가 형성되어있으며, 이형-이량체로 형성되어있다. B. Vac8-Vac17의 각각의 인터페이스. 인터페이스 I에서는 통해 Vac17의 메인체인과 Vac8의 아미노산 잔기를 통한 수소결합을 통해 형성된다. 인터페이스 II는 Vac17의 HC helix와 Vac8의 H1 helix의 소수성 결합을 통해 형성된다. |
|
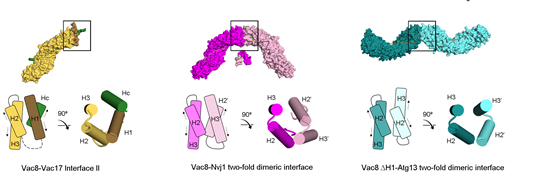
그림2. Vac8-Vac17, Vac8-Nvj1, Vac8-Atg13간의 4차 구조 비교.이형-이량체를 형성하는 Vac8-Vac17과 이형-사량체를 형성하는 Vac8-Nvj1, Vac8-Atg13 복합체. Vac17의 Hc helix를 통해 Vac8의 ARM1과 H1 helix가 묶이게 되어, Vac8-Vac17의 결합 인터페이스가 형성된다. 반면, Nvj1, Atg13이 각각 Vac8과 결합한 경우, H1 helix가 ARM1으로부터 떨어지게되어, Vac8간의 이량체 인터페이스가 형성이 되고, ARM1끼리 이량체를 형성한다. 그래서 Vac8-Nvj1, Vac8-Atg13이 각각 이형-사량체의 사차구조를 만들게 된다. |
|
그림3. Vac8-Vac17, Vac8-Nvj1, Vac8-Atg13 복합체 형성에 따른 서로다른 세포내 기작 조절.Vac8-Vac17 복합체가 형성이되면, Vac17의 Hc helix가 Vac8의 H1 helix를 붙잡아 동형-이량체 형성 인터페이스(ARM1)가 노출되는 것을 막게 된다. 이는 세포 내 자가포식을 일으키는 복합체 형성을 억제하고, Myo2-Actin과 함께 복합체를 형성해 딸세포로의 리소좀 수송을 진행시킨다. 반면, Nvj1과 Atg13이 각각 Vac8과 결합하게되면, H1 helix가 ARM1으로부터 분리가 되고, Vac8의 동형-이량체 형성 인터페이스가 드러나게 된다. 그래서 각각 Vac8-Nvj1, Vac8-Atg13 이형 사량체를 형성한다. 이렇게 형성된 4차 구조는 각각 아치형과 나선형 구조를 띄게되며, PMN, Cvt 과정을 통한 자가포식을 일으킨다. |
|
|
UNIST 홍보팀 news@unist.ac.kr TEL : 052)217-1230FAX : 052)217-1229 |