|
|
|
|
|
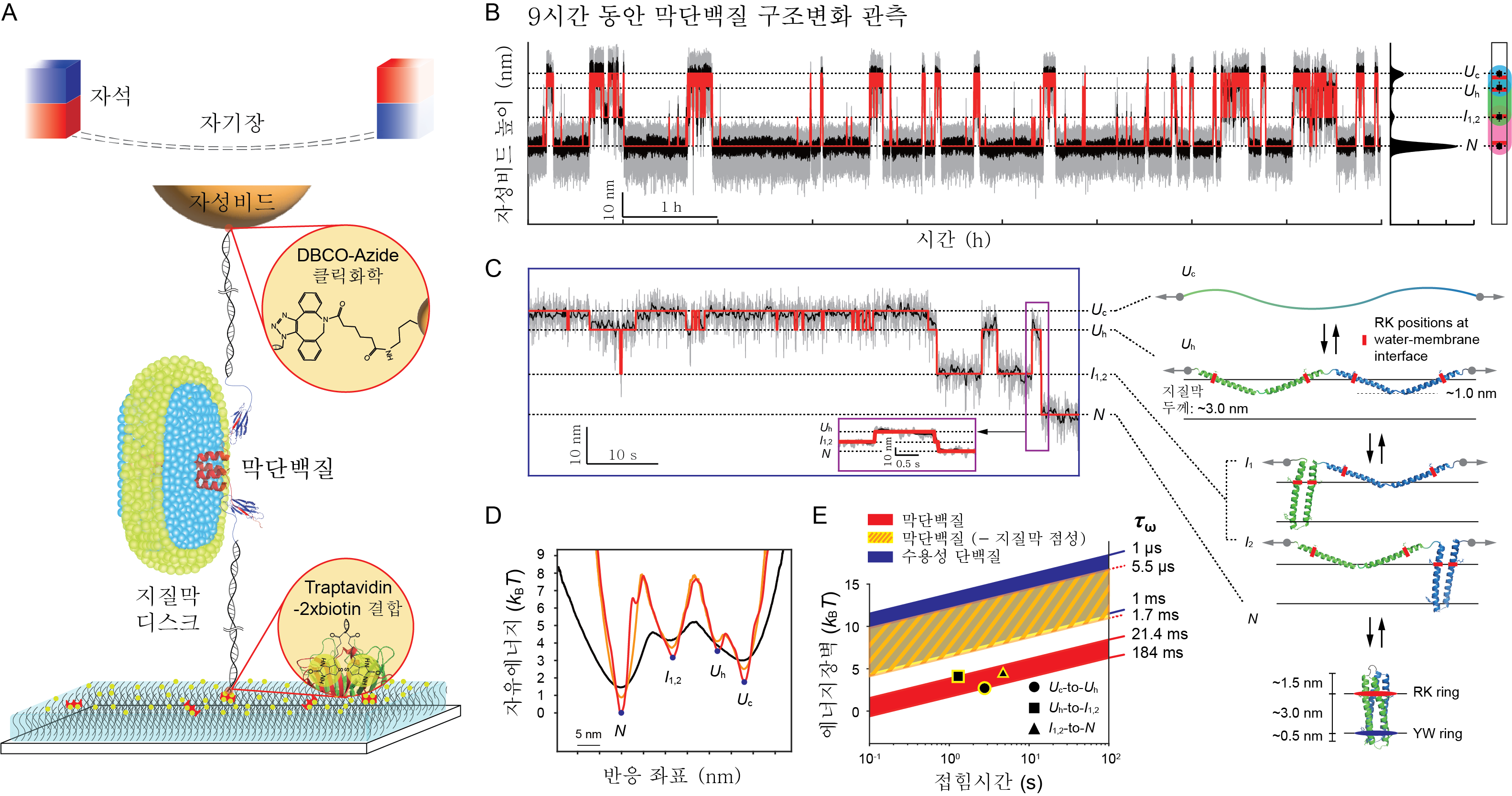
세포막 또는 세포 소기관의 막에 존재하는 단백질인 막단백질(membrane protein)은 세포간 신호전달, 면역반응 등에 관여하는 중요한 단백질 군이다. 시판되는 약물의 약 60% 정도가 막단백질을 표적으로 할 정도로 막단백질의 구조와 구조 형성(접힘, folding)의 원리를 이해하는 것은 의약학적으로 중요하다. UNIST(총장 이용훈) 화학과 민두영 교수팀은 막단백질이 구조를 형성하는데 걸리는 최소 시간인 “제한속도” 를 최초로 측정했다. 여러 연구를 통해 일반적인 수용성 단백질은 최대 1마이크로초(μs, 100만분의 1초) 정도의 제한속도를 갖는다고 알려져 있지만, 막단백질 접힘의 제한속도에 대해선 알려진 바가 없었다. 특히 접힘의 제한속도를 측정하기 위해선 막단백질의 구조가 빠르게 변화하는 전이(transition) 순간을 포착할 수 있어야 한다. 하지만 막단백질의 전이 현상은 매우 드물게 일어나고, 수십 피코뉴턴(pN, 1조분의 1뉴턴)의 작은 힘에서도 측정시스템이 쉽게 망가져 변화를 측정하기 어려웠다. 연구팀은 이러한 문제를 해결하기 위해 막단백질이 접히고 풀리는 구조 변화를 긴 시간 동안 관측할 수 있는 “튼튼한” 단일분자집게(single-molecule tweezers)를 개발했다. 개발된 단일분자집게를 활용해 12pN의 힘에서 9시간 동안 막단백질의 전이 현상을 관찰했다. 이렇게 최대 1,000번이 넘는 전이 발생의 순간을 포착해 다량의 데이터를 확보할 수 있었다. 연구팀은 관찰된 다량의 데이터를 통해 막단백질 구성의 최소단위인 나선헤어핀(helical hairpin)의 제한속도를 측정했다. 막단백질을 구성하는 나선헤어핀은 나선형 구조 2개가 헤어핀 형태로 결합돼 있는데, 이번 연구를 통해 접힘의 제한속도가 약 20ms(1천분의 1초) 임을 확인했다. 연구 결과 막단백질 접힘의 제한속도는 수용성 단백질에 비해 최대 2만 배 느린 속도이며, 이렇게 느린 속도는 단백질 응집(aggregation) 현상에 치명적일 수 있다는 결론을 도출했다. 또한 연구팀은 지질막의 점성이 나선형 구조 사이의 상호작용을 방해하기 때문에 이런 결과가 나타난다는 의견을 제시했다. 민두영 화학과 교수는 “보통 화학변화 또는 구조변화의 전이 순간과 관련된 제한속도는 측정하기가 쉽지 않지만, 이번 연구를 통해 막단백질 접힘의 제한속도를 최초로 규명했다”며 “본 연구결과는 막단백질 접힘 더 나아가 막단백질의 상호작용에 의미있는 통찰을 제공하며, 막단백질을 표적으로 하는 약물개발에 중요한 기초 지식으로 활용될 수 있을 것”이라고 전했다. 연구 결과는 생명과학분야 저명 학술지인 eLife에 5월 30일자로 온라인 게재됐다. 연구 수행은 과학기술정보통신부 한국연구재단 및 울산과학기술원의 지원을 받아 이뤄졌다. (논문명: Robust membrane protein tweezers reveal the folding speed limit of helical membrane proteins) |
|
|
|
[붙임] 연구결과 개요 |
1. 연구배경막단백질(membrane protein)의 구조, 구조형성(접힘), 기능을 이해하는 것은 막단백질을 표적으로 하는 약물개발에 중요하다. 실제로 시판되는 약물의 약 60% 정도가 막단백질을 표적으로 한다. 막단백질이 자연적으로 존재하는 환경은 지질막(lipid membrane)이고, 특정 지질분자는 막단백질의 구조적 변화를 유도하고 기능을 미세하게 조절하는 것으로 알려져 있다. 하지만 지질막을 세포밖에서 재구성해 막단백질의 구조변화를 포착하는 것은, 수용성 단백질 비해 상당한 기술적 어려움을 동반한다. 지질막 안에서의 막단백질 접힘 동역학은 최근에서야 연구되기 시작했으며, 막단백질의 구조가 얼마나 빠른 속도로 형성될 수 있는지는 알려진 바가 없다. 이는 접힘 “제한속도” 로 지칭되기도 하는데, 크래머속도이론(Kramers rate theory)에서 속도상수(rate constant)와 자유에너지장벽(free energy barrier)을 연결시키는 중요한 변수이다. 이전 연구에서는 수용성 단백질들에 대해 측정된 제한속도 값을 막단백질에 단순 적용하였으나, 본 연구에 따르면 최대 2 만배 만큼의 차이가 있음을 밝혔다. 이는 막단백질 접힘 동역학을 보다 정확히 이해하는 단초가 된다. |
2. 연구배경본 연구에서는 막단백질 구조형성의 “제한속도” 를 최초로 측정했다. 여기서 “제한속도” 란 단백질이 구조를 형성하는데 걸리는 최소 시간을 의미한다. 연구팀은 속도상수와 자유에너지장벽을 독립적으로 측정할 수 있다면, 크래머속도이론을 통해 막단백질 접힘 제한속도를 얻어낼 수 있음을 주목했다. 하지만 자유에너지장벽을 측정하기 위해서는 구조형성 전이(transition)가 발생하는 순간을 포착할 수 있어야 한다. 하지만 순간적인 전이 사건은 매우 드물게 일어나기 때문에 충분한 데이터를 얻기 어렵다. 따라서 매우 빠른 시간분해능을 이용하거나 매우 긴 시간동안 측정해 많은 양의 데이터를 확보해야 한다. 연구팀은 이러한 문제를 해결하기 위해, 막단백질이 접히고 풀리는 구조변화를 수 시간에 걸친 긴 시간동안 관측할 수 있는 “튼튼한” 단일분자집게(single-molecule tweezers)를 개발했다. 기존에 사용한 화학결합은 비공유결합으로 수십 pN 의 작은 힘에서도 분자결합이 떨어져 나가 시스템이 쉽게 무너지는 단점이 있었다. 본 연구에서는 DBCO 클릭화학(click chemistry)를 이용하여 공유결합으로 강하게 묶음으로써, 50 pN 의 높은 힘에서도 12시간의 긴 시간동안이나 시스템이 유지되는 것을 확인할 수 있었다. 연구팀이 개발한 분자집게 기법을 활용해, 최대 1000번이 넘는 구조변화를 관측하고 구조형성 전이가 발생하는 순간을 포착해 다량의 데이터를 확보했다. 측정된 데이터로부터 자유에너지장벽을 포함해 전체 자유에너지지평(free energy landscape)을 디컨볼루션(deconvolution) 방법을 이용해 정량하였다. 또한 접힘 속도상수는 측정 데이터를 은닉 마르코프 모델(hidden Markov model)로 분석해 추출했다. 연구팀은 나선헤어핀(helical hairpin) 구조형성의 제한속도가 약 20 ms 임을 도출해 낼 수 있었는데, 이는 비슷한 크기의 수용성 단백질에 비해 2 만배 정도나 느린 속도이다. 연구팀은 지질막(lipid membrane)의 점성(viscosity)이 나선구조간 상호작용을 방해하기 때문에 나타나는 현상이라고 제안했다. 이렇게 높은 점성을 띠는 용매 환경에서는 개개의 단백질보다 용매에 의한 마찰(friction) 효과가 접힘속도 결정에 지배적이다. 또한 막단백질의 약 80% 는 나선형(helix) 구조를 가지고, 이러한 막단백질의 3차 구조 접힘의 최소단위는 나선헤어핀 구조로 알려져 있다. 이는 나선형 막단백질의 접힘 속도는 약 20 ms 의 시간스케일에 의해 제한됨을 시사한다. |
3. 기대효과본 연구에서 측정된 막단백질 접힘 제한속도 결과는 기존에 측정된 접힘 자유에너지 값들이 상당한 오류가 있음을 시사한다. 또한 지질막의 여러 물리화학적 특성 중 점성이 막단백질 접힘에 중요한 요인으로 작용할 수 있음을 의미한다. 본 연구는 막단백질 접힘 더 나아가 막단백질 이합체형성(dimerization)과 같은 상호작용에 의미있는 통찰을 제공하며, 막단백질을 표적으로 하는 약물개발에 기반지식으로 활용될 수 있을 것으로 기대된다. |
|
[붙임] 용어설명 |
1. 막단백질(membrane protein)세포막 또는 세포 소기관의 막에 존재하는 단백질. 세포간 신호전달, 면역반응, 이온항상성 등에 관여하는 생명현상에서 중요한 위치를 차지하는 단백질군. |
2. 지질막(lipid membrane)세포 및 세포 소기관의 내부와 외부를 구분 짓는 지질분자로 이루어진 2차원 막. 지질막은 막단백질과의 상호작용을 통해 막단백질의 구조와 기능을 조절한다고 알려져 있음. |
3. 크래머속도이론(Kramers rate theory)1940년에 네덜란드 과학자 핸드릭 크레머가 제안한 이론으로 화학변화가 일어날 때의 반응속도와 에너지장벽간의 관계식으로 요약됨. |
4. 단일분자집게(single-molecule tweezers)단일분자에 수~수십 pN 의 힘을 가해 구조변화를 nm(10억분의 1미터) 수준의 공간분해능으로 관측할 수 있는 기술. 대표적으로 자기집게와 광학집게가 있음. |
5. 자유에너지지평(free energy landscape)반응좌표에 따른 자유에너지 변화를 나타낸 그래프 |
6. 디컨볼루션(deconvolution)컨볼루션(convolution)의 역과정. 측정 과정에서 컨볼루션의 형태로 외부신호가 섞여 있을 때 원본신호를 추출하는 과정. |
7. 은닉 마르코프 모델(hidden Markov model)은닉된 상태와 관찰가능한 결과인 두 가지 요소로 이루어진 마르코프 모델. 여기서는 측정 신호로부터 구조적 상태를 추정할 때 사용. |
8. 나선헤어핀(helical hairpin)단백질의 2차 구조인 나선(helix)구조 2개가 헤어핀 형태로 결합된 3차 구조. 나선형 막단백질 3차 구조의 최소 단위. |
|
[붙임] 그림설명 |
|
그림1. “튼튼한”단일분자집게를 이용한 막단백질 구조형성 및 제한속도 관측.(A) DBCO 클릭화학 및 traptavidin 결합을 이용한 단일분자 자기집게 기법. (B) 12pN의 힘에서 9시간의 긴 시간 동안 막단백질 구조변화 측정. (C) 측정신호를 중합체(polymer) 모델을 이용해 분석한 막단백질 구조 상태들. (D) 막단백질 접힘 자유에너지지평. (E) 접힘 제한속도에 따른 에너지장벽-접힘시간 평면. 제한속도 차이에 의해 막단백질과 수용성 단백질에 해당하는 영역이 상당한 격차를 보임. |
|
|
UNIST 홍보팀 news@unist.ac.kr TEL : 052)217-1230FAX : 052)217-1229 |





![[연구그림] “튼튼한”단일분자집게를 이용한 막단백질 구조형성 및 제한속도 관측](https://news.unist.ac.kr/kor/wp-content/uploads/2014/11/press_20141125_03.jpg)